Стадии мейоза. Мейоз. Особенности первого и второго деления мейоза. Биологическое значение мейоза. Отличия мейоза от митоза Биологическое значение мейоза
Клетка размножается путем деления. Существуют два способа деления: митоз и мейоз.
Митоз (от греч. митос - нитка), или непрямое деление клетки, представляет собой непрерывный процесс, в результате которого происходит сначала удвоение, а затем равномерное распределение наследственного материала, содержащегося в хромосомах, между двумя образующимися клетками. В этом его биологическое значение. Деление ядра влечет за собой деление всей клетки. Этот процесс называется цитокинезом (от греч. цитос - клетка).
Состояние клетки между двумя митозами называют интерфазой, или интеркинезом, а все происходящие в ней во время подготовки к митозу и в период деления изменения - митотическим, или клеточным, циклом.
У разных клеток митотические циклы имеют разную продолжительность. Большую часть времени клетка находится в состоянии интеркинеза, митоз длится сравнительно недолго. В общем митотическом цикле собственно митоз занимает 1/25-1/20 времени, и у большинства клеток он продолжается от 0,5 до 2 ч.
Толщина хромосом столь мала, что при рассмотрении интерфазного ядра в световой микроскоп они не видны, удается лишь различить гранулы хроматина в узлах их скручивания. Электронный микроскоп позволил обнаруживать хромосомы и в неделящемся ядре, хотя они в это время очень длинны и состоят из двух нитей хроматид, диаметр каждой из которых составляет всего 0,01 мкм. Следовательно, хромосомы в ядре не исчезают, а принимают форму длинных и тонких нитей, которые почти не видны.
Во время митоза ядро проходит четыре последовательные фазы: профазу, метафазу, анафазу и телофазу.
Профаза (от греч. про - раньше, фазис - проявление). Это первая фаза деления ядра, во время которой внутри ядра появляются структурные элементы, имеющие вид тонких двойных нитей, что и обусловило название этого типа деления - митоз. В результате спирализации хромонем хромосомы в профазе уплотняются, укорачиваются и становятся отчетливо видимыми. К концу профазы можно хорошо наблюдать, что каждая хромосома состоит из двух тесно соприкасающихся одна с другой хроматид. В дальнейшем обе хроматиды соединяются общим участком - центромерой и начинают постепенно передвигаться к клеточному экватору.
В середине или в конце профазы ядерная оболочка и ядрышки исчезают, центриоли удваиваются и отходят к полюсам. Из материала цитоплазмы и ядра начинает формироваться веретено деления. Оно состоит из двух видов нитей: опорных и тянущих (хромосомных). Опорные нити составляют основу веретена, они тянутся от одного полюса клетки к другому. Тянущие нити соединяют центромеры хроматид с полюсами клетки и обеспечивают в последующем движение к ним хромосом. Митотический аппарат клетки очень чувствителен к различным внешним воздействиям. При действии радиации, химических веществ и высокой температуры клеточное веретено может разрушаться, возникают всевозможные неправильности в делении клетки.
Метафаза (от греч. мета - после, фазис - проявление). В метафазе хромосомы сильно уплотняются и приобретают определенную, характерную для данного вида форму. Дочерние хроматиды в каждой паре разъединены хорошо видимой продольной щелью. Большинство хромосом становится двуплечими. Местом перегиба - центромерой - они прикрепляются к нити веретена. Все хромосомы располагаются в экваториальной плоскости клетки, свободные концы их направлены к центру клетки. В это время хромосомы лучше всего наблюдать и подсчитывать. Очень отчетливо видно и клеточное веретено.
Анафаза (от греч. ана - вверх, фазис - проявление). В анафазе вслед за делением центромер начинается расхождение хроматид, ставших теперь отдельными хромосомами, к противоположным полюсам. При этом хромосомы имеют вид разнообразных крючков, обращенных своими концами к центру клетки. Так как из каждой хромосомы возникли две совершенно одинаковые хроматиды, то в обеих образовавшихся дочерних клетках число хромосом будет равно диплоидному числу исходной материнской клетки.
Процесс деления центромер и движения к разным полюсам всех вновь образовавшихся парных хромосом отличается исключительной синхронностью.
В конце анафазы начинается раскручивание хромонемных нитей, и хромосомы, отошедшие к полюсам, видны уже не так четко.
Телофаза (от греч. телос - конец, фазис - проявление). В телофазе продолжается деспирализация хромосомных нитей, и хромосомы постепенно становятся более тонкими и длинными, приближаясь к тому состоянию, в котором они были в профазе. Вокруг каждой группы хромосом образуется ядерная оболочка, формируется ядрышко. В это же время завершается деление цитоплазмы и возникает клеточная перегородка. Обе новые дочерние клетки вступают в период интерфазы.
Весь процесс митоза, как уже отмечалось, занимает не более 2 ч. Продолжительность его зависит от вида и возраста клеток, а также от внешних условий, в которых они находятся (температура, освещенность, влажность воздуха и т. д.). Отрицательно сказываются на нормальном ходе деления клеток высокие температуры, радиация, различные наркотики и растительные яды (колхицин, аценафтен и др.).
Митотическое деление клеток отличается высокой степенью точности и совершенства. Механизм митоза создавался и совершенствовался на протяжении многих миллионов лет эволюционного развития организмов. В митозе находит свое проявление одно из важнейших свойств клетки как самоуправляемой и, самовоспроизводящейся живой биологической системы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Мейоз – это особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в два раза (от древнегреч. «мейон» – меньше – и от «мейозис» – уменьшение).
Главной особенностью мейоза является конъюгация (спаривание) гомологичных хромосом с последующим расхождением их в разные клетки. Поэтому в первом делении мейоза вследствие образования бивалентов к полюсам клетки расходятся не однохроматидные, а двухроматидные хромосомы. В результате число хромосом уменьшается в два раза, и из диплоидной клетки образуются гаплоидные клетки.
Исходное число хромосом в клетке, которая вступает в мейоз, называется диплоидным (2n). Число хромосом в клетках, образовавшихся в ходе мейоза, называется гаплоидным (n).
Мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому его называют редукционным. Во втором делении число хромосом не изменяется; поэтому его называют эквационным (уравнивающим).
Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца: примерно 0,2...0,4 % ДНК остается неудвоенной. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с. При наличии центриолей происходит их удвоение. Таким образом, в клетке имеется две диплосомы, каждая из которых содержит пару центриолей.
Первое деление мейоза (редукционное, или мейоз I)
Сущность редукционного деления заключается в уменьшении числа хромосом в два раза: из исходной диплоидной клетки образуется две гаплоидные клетки с двухроматидными хромосомами (в состав каждой хромосомы входит 2 хроматиды).
Профаза I (профаза первого деления) включает ряд стадий.
Лептотена (стадия тонких нитей). Хромосомы видны в световой микроскоп в виде клубка тонких нитей.
Зиготена (стадия сливающихся нитей). Происходит конъюгация гомологичных хромосом (от лат. conjugatio – соединение, спаривание, временное слияние). Гомологичные хромосомы (или гомологи) – это парные хромосомы, сходные между собой в морфологическом и генетическом отношении. В результате конъюгации образуются биваленты. Бивалент – это относительно устойчивый комплекс из двух гомологичных хромосом. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов. Количество бивалентов равно гаплоидному числу хромосом. Иначе биваленты называются тетрады, так как в состав каждого бивалента входит 4 хроматиды.
Пахитена (стадия толстых нитей). Хромосомы спирализуются, хорошо видна их продольная неоднородность. Завершается репликация ДНК. Завершается кроссинговер – перекрест хромосом, в результате которого они обмениваются участками хроматид.
Диплотена (стадия двойных нитей). Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных точках, которые называются хиазмы (от древнегреч. буквы χ – «хи»).
Диакинез (стадия расхождения бивалентов). Хиазмы перемещаются к теломерным участкам хромосом. Биваленты располагаются на периферии ядра. В конце профазы I ядерная оболочка разрушается, и биваленты выходят в цитоплазму.
Метафаза I (метафаза первого деления). Формируется веретено деления. Биваленты перемещаются в экваториальную плоскость клетки. Образуется метафазная пластинка из бивалентов.
Анафаза I (анафаза первого деления). Гомологичные хромосомы, входящие в состав каждого бивалента, разъединяются, и каждая хромосома движется в сторону ближайшего полюса клетки. Разъединения хромосом на хроматиды не происходит.
Телофаза I (телофаза первого деления). Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. В норме каждая дочерняя клетка получает одну гомологичную хромосому из каждой пары гомологов. Формируются два гаплоидных ядра, которые содержат в два раза меньше хромосом, чем ядро исходной диплоидной клетки. Каждое гаплоидное ядро содержит только один хромосомный набор, то есть каждая хромосома представлена только одним гомологом. Содержание ДНК в дочерних клетках составляет 2с.
В большинстве случаев (но не всегда) телофаза I сопровождается цитокинезом.
После первого деления мейоза наступает интеркинез – короткий промежуток между двумя мейотическими делениями. Интеркинез отличается от интерфазы тем, что не происходит репликации ДНК, удвоения хромосом и удвоения центриолей: эти процессы произошли в предмейотической интерфазе и, частично, в профазе I.
Второе деление мейоза (эквационное, или мейоз II)
В ходе второго деления мейоза уменьшения числа хромосом не происходит. Сущность эквационного деления заключается в образовании четырех гаплоидных клеток с однохроматидными хромосомами (в состав каждой хромосомы входит одна хроматида).
Профаза II (профаза второго деления). Не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей. В каждой из дочерних клеток формируется веретено деления.
Метафаза II (метафаза второго деления). Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток независимо друг от друга. Эти экваториальные плоскости могут быть параллельны друг другу или взаимно перпендикулярны.
Анафаза II (анафаза второго деления). Хромосомы разделяются на хроматиды (как при митозе). Получившиеся однохроматидные хромосомы в составе анафазных групп перемещаются к полюсам клеток.
Телофаза II (телофаза второго деления). Однохроматидные хромосомы полностью переместились к полюсам клетки, формируются ядра. Содержание ДНК в каждой из клеток становится минимальным и составляет 1с.
Таким образом, в результате описанной схемы мейоза из одной диплоидной клетки образуется четыре гаплоидные клетки. Дальнейшая судьба этих клеток зависит от таксономической принадлежности организмов, от пола особи и ряда других факторов.
Типы мейоза. При зиготном и споровом мейозе образовавшиеся гаплоидные клетки дают начало спорам (зооспорам). Эти типы мейоза характерны для низших эукариот, грибов и растений. Зиготный и споровый мейоз тесно связан со спорогенезом. При гаметном мейозе из образовавшихся гаплоидных клеток образуются гаметы. Этот тип мейоза характерен для животных. Гаметный мейоз тесно связан с гаметогенезом и оплодотворением. Таким образом, мейоз – это цитологическая основа полового и бесполого (спорового) размножения.
Биологическое значение мейоза. Немецкий биолог Август Вайсман (1887) теоретически обосновал необходимость мейоза как механизма поддержания постоянного числа хромосом. Поскольку при оплодотворении ядра половых клеток сливаются (и, тем самым, в одном ядре объединяются хромосомы этих ядер), и поскольку число хромосом в соматических клетках остается константным, то постоянному удвоению числа хромосом при последовательных оплодотворениях должен противостоять процесс, приводящий к сокращению их числа в гаметах ровно вдвое. Таким образом, биологическое значение мейоза заключается в поддержании постоянства числа хромосом при наличии полового процесса. Мейоз обеспечивает также комбинативную изменчивость – появление новых сочетаний наследственных задатков при дальнейшем оплодотворении.
Деление клеток посредством мейоза проходит в два основных этапа: мейоз I и мейоз II. В конце мейотического процесса образуются четыре . Прежде чем делящаяся клетка попадет в мейоз, она проходит через период , называемый интерфазой.
Интерфаза
- Фаза G1: этап развития клетки перед синтезом ДНК. На этой стадии клетка подготавливаясь к делению увеличивается в массе.
- S-фаза: период, в течение которого синтезируется ДНК. Для большинства клеток эта фаза занимает короткий промежуток времени.
- Фаза G2: период после синтеза ДНК, но до начала профазы. Клетка продолжает синтезировать дополнительные белки и увеличиваться в размерах.
В последней фазе интерфазы клетка все еще имеет нуклеолы. окружено ядерной мембраной, а клеточное хромосомы дублируются, но находятся в форме . В две пары , образованных из репликации одной пары, расположены за пределами ядра. В конце интерфазы клетка переходит в первый этап мейоза.
Мейоз I:
Профаза I
В профазе I мейоза происходят следующие изменения:
- Хромосомы конденсируются и присоединяются к ядерной оболочке.
- Возникает синапсис (попарное сближение гомологичных хромосом) и образуется тетрада. Каждая тетрада состоит из четырех хроматид.
- Может произойти генетическая рекомбинация.
- Хромосомы сгущаются и отсоединяются от ядерной оболочки.
- Подобно , центриоли мигрируют друг от друга, а ядерная оболочка и ядрышки разрушаются.
- Хромосомы начинают миграцию к метафазной (экваториальной) пластине.
В конце профазы I клетка входит в метафазу I.
Метафаза I
 В метафазе I мейоза происходят следующие изменения:
В метафазе I мейоза происходят следующие изменения:
- Тетрады выравниваются на метафазной пластине.
- гомологичных хромосом ориентированы на противоположные полюса клетки.
В конце метафазы I клетка входит в анафазу I.
Анафаза I
 В анафазе I мейоза происходят происходят следующие изменения:
В анафазе I мейоза происходят происходят следующие изменения:
- Хромосомы перемещаются в противоположные концы клетки. Подобно митозу, кинетохоры взаимодействуют с микротрубочками, чтобы переместить хромосомы к полюсам клетки.
- В отличие от митоза, остаются вместе после того, как перемещаются в противоположные полюса.
В конце анафазы I клетка входит в телофазу I.
Телофаза I
 В телофазе I мейоза происходят следующие изменения:
В телофазе I мейоза происходят следующие изменения:
- Волокна веретена продолжают перемещать гомологичные хромосомы на полюса.
- Как только движение завершено, каждый полюс клетки имеет гаплоидное количество хромосом.
- В большинстве случаев цитокинез (деление ) происходит одновременно с телофазой I.
- В конце телофазы I и цитокинеза образуются две дочерние клетки, каждая из которых имеет половину числа хромосом исходной родительской клетки.
- В зависимости от типа клетки могут возникать различные процессы при подготовке к мейозу II. Однако генетический материал не реплицируется снова.
В конце телофазы I клетка входит в профазу II.
Мейоз II:
Профаза II
 В профазе II мейоза происходят следующие изменения:
В профазе II мейоза происходят следующие изменения:
- Ядерная и ядра разрушаются, пока появляется веретено деления.
- Хромосомы больше не реплицируются в этой фазе.
- Хромосомы начинают мигрировать к метафазной пластинке II (на экваторе клеток).
В конце профазы II клетки входят в метафазу II.
Метафаза II
 В метафазе II мейоза происходят следующие изменения:
В метафазе II мейоза происходят следующие изменения:
- Хромосомы выстраиваются на метафазной пластинке II в центре клеток.
- Кинетохорные нити сестринских хроматид расходятся к противоположным полюсам.
В конце метафазы II клетки входят в анафазу II.
Анафаза II
 В анафазе II мейоза происходят следующие изменения:
В анафазе II мейоза происходят следующие изменения:
- Сестринские хроматиды разделяются и начинают перемещаться к противоположным концам (полюсам) клетки. Волокна веретена деления, не связанные с хроматидами, вытягиваются и удлиняют клетки.
- Как только парные сестринские хроматиды отделены друг от друга, каждая из них считается полной хромосомой, называемые .
- При подготовке к следующему этапу мейоза два полюса клеток также отдаляются друг от друга во время анафазы II. В конце анафазы II каждый полюс содержит полную компиляцию хромосом.
После анафазы II клетки входят в телофазу II.
Телофаза II
 В телофазе II мейоза происходят следующие изменения:
В телофазе II мейоза происходят следующие изменения:
- Образуются отдельные ядра на противоположных полюсах.
- Происходит цитокинез (деление цитоплазмы и образование новых клеток).
- В конце мейоза II производятся четыре дочерние клетки. Каждая клетка имеет половину числа хромосом от исходной родительской клетки.
Результат мейоза
Конечным результатом мейоза является производство четырех дочерних клеток. Эти клетки имеют в двое меньше хромосом относительно родительской. При мейозе продуцируются только половые . Другие делятся посредством митоза. Когда половые объединяются во время оплодотворения, они становятся . Диплоидные клетки имеют полный набор гомологичных хромосом.
Профаза 2 (1n2c ). Короче профазы 1, хроматин конденсирован, нет конъюгации и кроссинговера, происходят процессы, обычные для профазы – распад ядерных мембран на фрагменты, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления.
Метафаза 2
(1n2c
). Двухроматидные хромосомы выстраиваются в экваториальной плоскости клетки, формируется метафазная пластинка.
Создаются предпосылки для третьей рекомбинации генетического материала – многие хроматиды мозаичные и от их расположения на экваторе зависит, к какому полюсу они в дальнейшем отойдут. К центромерам хроматид прикрепляются нити веретена деления.
Анафаза 2 (2n2с). Происходит деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами),происходит третья рекомбинация генетического материала.
Телофаза 2 (1n1c в каждой клетке). Хромосомы деконденсируются, образуются ядерные оболочки, разрушаются нити веретена деления, появляются ядрышки, происходит деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток.
5. Отличие мейоза I от мейоза II
1.Первому делению предшествует ннтерфаза с редупликацией хромомом, при втором делении редпликации генетического материала нет, то есть отсутствует синтетическая стадия.
2.Профаза первого деления длительная.
3.В первом делении происходит конъюгация хромосом и
кроссинговер.
4.В первом делении к полюсам расходятся гомологичные хромосомы (биваленты, состоящие из пары хроматид), а во втором – хроматиды.
Мейоз: 1 - лептотена; 2 - зиготена; 3 - пахитена; 4 - диплотена; 5 - диакинез; 6 - метафаза 1; 7 - анафаза 1; 8 - телофаза 1; 9 - профаза 2; 10 - метафаза 2; 11 - анафаза 2; 12 - телофаза 2.
6. Отличия мейоза от митоза
1. В митозе одно деление, а в мейозе – два (из-за этого получается 4 клетки).
2. В профазе первого деления мейоза происходит конъюгация (тесное сближение гомологичных хромосом) и кроссинговер (обмен участками гомологичных хромосом), это приводит к перекомбинации (рекомбинации) наследственной информации.
3. В анафазе первого деления мейоза происходит независимое расхождение гомологичных хромосом (к полюсам клетки расходятся двуххроматидные хромосомы). Это приводит к рекомбинации и редукции.
4. В интерфазе между двумя делениями мейоза удвоения хромосом не происходит, поскольку они и так двойные.
5. После митоза получается две клетки, а после мейоза – четыре.
6. После митоза получаются соматические клетки (клетки тела), а после мейоза – половые клетки (гаметы – сперматозоиды и яйцеклетки; у растений после мейоза получаются споры).
7. После митоза получаются одинаковые клетки (копии), а после мейоза – разные (происходит рекомбинация наследственной информации).
8. После митоза количество хромосом в дочерних клетках остается таким же, как было в материнской, а после мейоза уменьшается в 2 раза (происходит редукция числа хромосом; если бы её не было, то после каждого оплодотворения число хромосом возрастало бы в два раза; чередование редукции и оплодотворения обеспечивает постоянство числа хромосом).
7. Биологическое значение мейоза
Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. С его помощью поддерживается постоянство хромосомного набора – после слияния гамет не происходит его удвоения. Благодаря мейозу образуются генетически различные клетки, т.к. в процессе мейоза трижды происходит перекомбинация генетического материала: за счет кроссинговера (профаза 1), за счет случайного, независимого расхождения гомологичных хромосом (анафаза 1) и за счет случайного расхождения хроматид (анафаза 2).
8. Способы размножения организмов
9. Отличие полового размножения от бесполого

10. Основные формы бесполого размножения: деление на два (митоз), множественное деление (шизогония), почкование, фрагментация, спорообразование, вегетативное размножение, полиэмбриония).
Бесполое размножение – процесс возникновения дочерних особей из одно или группы соматических клеток материнского организма. Этот способ размножения более древний. В его основе лежит митотическое деление клеток. Значение бесполого размножения заключается в быстром увеличении числа особей, почти не различающихся между собой. Различают следующие формы бесполого размножения:
1.Деление надвое – приводит к возникновению из одного родительского организма двух дочерних. Является преобладающей формой деления у прокариот и простейших. Различные одноклеточные животные делятся по-разному. Так, жгутиковые делятся продольно, а инфузории – поперечно. Такое деление встречается и у многоклеточных животных – кишечнополостных (продольное деление у медуз) и червей (поперечное деление у кольчатых червей).
3.Почкование – на теле материнского организма возникает скопление клеток, которое растет и постепенно приобретает сходство с материнской особью. Затем дочерняя особь отделяется и начинает вести самостоятельное существование. Такое размножение распространено среди низших многоклеточных (губки, кишечнополостные, мшанки, некоторые черви и оболочники). Иногда дочерние особи не отделяются полностью от родительской, что приводит к образованию колоний.
4.Фрагментация – происходит распад тела многоклеточного организма на части, которые в дальнейшем превращаются в самостоятельные особи (плоские черви, иглокожие).
5.Спорами – дочерний организм развивается из специализированной клетки-споры.
Различают две основные формы бесполого размножения растений: вегетативное размножение и спорообразование. Вегетативное размножение одноклеточных растений осуществляется простым делением одной клетки на две. У грибов формы его более разнообразны – спорообразование (плесневые грибы, шляпочные) и почкование (дрожжи). У покрытосеменных растений вегетативное размножение происходит за счет вегетативных (неполовых) органов – корня, стебля, листа.
У некоторых видов животных наблюдается полиэмбриония – бесполое размножение зародыша, образовавшегося путем полового размножения. Например, у броненосцев на стадии бластулы происходит разделение клеточного материала первоначально одного зародыша между 4–8 зародышами, из которых в последствии развиваются полноценные особи. В результате полиэмбрионии у человека рождаются однояйцовые близнецы.

11. Основные формы полового размножения у одноклеточных организмов (конъюгация, копуляция) и у многоклеточных организмов (без оплодотворения (партеногенез) и с оплодотворением).
Половое размножение – наблюдается в жизненных циклах всех основных групп организмов. Распространенность полового размножения объясняется тем, что оно обеспечивает значительное генетическое разнообразие и, следовательно, фенотипическую изменчивость потомства.
В основе полового размножения лежит половой процесс, суть которого сводится к объединению в наследственном материале для развития потомка генетической информации от двух разных источников – родителей.
Одной из форм полового процесса является конъюгация. При этом происходит временное соединение двух особей с целью обмена (рекомбинации) наследственным материалом, например, у инфузорий. В результате появляются особи генетически отличные от родительских организмов, которые в дальнейшем осуществляют бесполое размножение. Число инфузорий после конъюгации не изменяется, поэтому говорить в прямом смысле о размножении в этом случае нельзя.
У простейших половой процесс может осуществляться и в форме копуляции – слияния двух особей в одну, объединение и рекомбинация наследственного материала. Далее такая особь размножается делением.
Для участия в половом размножении в родительских организмах вы-рабатываются гаметы – клетки, специализированные к обеспечению генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы – клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития.
У некоторых организмов зигота образуется в результате объединения гамет, которые не отличаются по строению – явление изогамии. У большинства же видов половые клетки по структурным и функциональным признакам делятся на материнские (яйцеклетки) и отцовские (сперматозоиды).
Иногда развитие дочернего организма происходит из неоплодотворенной яйцеклетки. Это явление называют девственным развитием или партеногенезом. Источником наследственного материала для развития потомка в этом случае обычно служит ДНК яйцеклетки – явление гиногенеза. Реже наблюдается андрогенез – развитие потомка из клетки с цитоплазмой ооцита и ядром сперматозоида. Ядро женской гаметы в случае андрогенеза погибает.

12. Биологическое значение полового размножения
На определенном этапе эволюции у многоклеточных организмов половой процесс как способ обмена генетической информацией между особями в пределах вида оказался связанным с размножением. При половом размножении возникающие новые особи обычно отличаются от родительских и друг от друга комбинацией аллелей генов. Новые сочетания хромосом и генов проявляются у потомков новым сочетанием признаков. В результате возникает большое разнообразие особей в пределах одного вида. Таким образом, биологическое значение полового размножения заключается не только в самовоспроизведении, но и в обеспечении исторического развития видов, то есть жизни как таковой. Это позволяет считать половое размножение биологически более прогрессивным, чем бесполое.
13. Сперматогенез
Процесс образования мужских половых клеток – сперматогенез. В результате образуются сперматозоиды.
В сперматогенезе различают 4 периода: размножение, рост, созревание (мейоз) и формирование (рис. 3).
В период размножения исходные недифференцированные половые клетки сперматогонии , или гонии делятся путем обычного митоза. Проделав несколько таких делений, они вступают в период роста. На этой стадии их называют сперматоцитами I порядка (или цитами I ). Они усиленно ассимилируют питательные вещества, укрупняются, претерпевают глубокую физико-химическую перестройку, в результате которой подготавливаются к третьему периоду – созреванию, или мейозу .
В мейозе сперматоциты I проходят два процесса клеточного деления. В первом делении (редукционном) происходит уменьшение числа хромосом (редукция). В результате из одного цита I возникает две равновеликие клетки – сперматоциты II порядка, или циты II. Затем наступает второе деление созревания. Оно протекает как обычный соматический митоз, но при гаплоидном числе хромосом. Такое деление называется эквационным («эквацио» – равенство), так как образуются две тождественные, т.е. полностью равноценные клетки, которые называются сперматидами.
В четвертом периоде – формирования – округлая сперматида приобретает форму зрелой мужской половой клетки: у нее вырастает жгутик, уплотняется ядро, образуется оболочка. В результате всего процесса сперматогенеза из каждой исходной недифференцированной сперматогонии получается 4 зрелых половых клетки, содержащих по гаплоидному набору хромосом.
На рис. 4 представлена схема процессов сперматогенеза и спермиогенеза у человека. Сперматогенез происходит в извитых семенных канальцах семенников.Развитие сперматозоидов начинается в период пренатального развития при закладке генеративных тканей, затем возобновляется в период наступления половозрелости и продолжается до старости.

Мужские половые клетки не развиваются одиночно, они растут в клонах и объединены между собой цитоплазматическими мостиками. Цитоплазматические мостики имеются между сперматогониями, сперматоцитами и сперматидами. В конце фазы формирования сперматозоиды освобождаются от цитоплазматических мостиков. У человека максимум дневной продуктивности сперматозоидов 108, продолжительность существования сперматозоида во влагалище до 2,5 ч, а в шейке матки до 48 ч.

14. Овогенез. Понятие о менструальном цикле
Процесс развития женских половых клеток называется овогенезом (оогенезом).
В овогенезе различают 3 периода: размножение, рост и созревание.
Недифференцированные женские половые клетки – овогонии – размножаются так же, как и сперматогонии, путем обычного митоза.
После деления они становятся овоцитами I порядка и переходят в период роста. Рост овоцитов длится очень долго – недели, месяцы и даже годы.
Затем овоцит I порядка вступает в период созревания, или мейоз. Здесь тоже совершаются редукционное и эквационное деления. Процессы деления в ядре протекают так же, как при мейозе сперматоцитов, но судьба цитоплазмы совершенно иная. При редукционном делении одно ядро увлекает с собой бульшую часть цитоплазмы , а на долю другого остается лишь незначительная ее часть. Поэтому образуется только одна полноценная клетка – овоцит II порядка, и вторая крошечная – направительное, или редукционное, тельце, которое может делиться на два редукционных тельца.
При втором, эквационном делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная клетка – овотида и третье полярное тельце. Овотида по составу ядра и функционально является вполне зрелой половой клеткой.
Период формирования, в отличие от сперматогенеза, в овогенезе отсутствует.
Таким образом, в овогенезе из одной овогонии возникает только одна зрелая яйцеклетка. Полярные тельца остаются недоразвитыми и вскоре погибают и фагоцитируются другими клетками. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду – икрой.
Развитие женских половых клеток происходит в яичниках. Период размно-жения наступает у оогоний еще у зародыша и прекращается к моменту рождения девочки.

Период роста при оогенезе более продолжительный, т.к. кроме подготовки к мейозу осуществляется накопление запаса питательных веществ, которые будут необходимы в дальнейшем для первых дроблений зиготы. В фазе малого роста происходит образование большого количества разных типов РНК.
В период большого роста фолликулярные клетки яичника образуют несколько слоев вокруг ооцита I порядка, что способствует переносу питательных веществ, синтезированных в других местах, в цитоплазму ооцита.
У человека период роста ооцитов может составлять 12–50 лет. После завершения периода роста ооцит I порядка вступает в период созревания.
В результате при оогенезе получается 4 клетки, из которых только одна станет в дальнейшем яйцеклеткой, а остальные 3 (полярные тельца) редуцируются. Биологическая значимость этого этапа оогенеза – сохранить все накопленные вещества цитоплазмы около одного гаплоидного ядра для обеспечения нормального питания и развития оплодотворенной яйцеклетки.
При оогенезе у женщин на стадии второй метафазы образуется блок, который снимается во время оплодотворения, и фаза созревания заканчивается только после проникновения сперматозоида в яйцеклетку.
Процесс оогенеза у женщин – это циклический процесс, повторяющийся примерно через каждые 28 дней (начиная с периода роста и заканчивая период только после оплодотворения). Этот цикл называется менструальным.
Отличительные особенности сперматогенеза и овогенеза у человека представлены в таблице 3.

Чем стадии мейоза отличаются от стадий митоза?
Основные отличия перечислены на схеме ниже. Но в действительности их намного больше. В мейозе два стадии - мейоз 1 и мейоз 2. В мейозе по-другому изменяется набор хромосом и молекул ДНК внутри стадий. Мейоз 2 похож на митоз в анафазе 2.
Рисунок 1. Отличия митоза от мейоза
Зачем существует профаза 1 мейоза 1? Какие ей можно дать метафоры?
Смысл существования профазы 1 - разнообразие жизни на Земле, так как в ней идет кроссинговер. Более того, любая профаза (митоза и мейоза) - это великий разрушитель и созидатель одновременно. Как разрушитель она выступает при растворении ядерной оболочки и ядрышка. Как созидатель - при создании видимых двухроматидных хромосом. Созидательная сила профазы проявляется также в разросшихся микротрубочках веретена деления и в отчетливом появлении двух полюсов деления клетки.
Что такое хроматиды? Чем они отличаются от хромосом?
В конце профазы конденсация хромосом завершается. Хромосомы утолщены, отделены от ядерной мембраны. В профазе становятся видны хромосомы, состоящие из двух хроматид. Представьте, что пара рук человека - это одна хромосома. В профазе мы четко видим, что одна хромосома состоит из двух частей - двух хроматид, точно также как человек имеет две руки, правую и левую.
Что представляют собой гомологичные хромосомы в профазе?
Гомологичные хромосомы, образно говоря - это муж и жена или мужчина и женщина. Почему? Во-первых, они парные, то есть находятся рядом друг с другом. Во-вторых, они достаются организму от разных родителей, всегда разнополых. В-третьих, в этой паре хромосом содержатся два аллеля. Они отвечают за альтернативные проявления одного гена. Например, есть ген цвета волос, и он представлен двумя аллелями: светлых волос и темных волос. Хромосомы в профазе - это гении общения. Они действительно «общаются», обмениваясь участками, в которых расположены определенные аллели. Следовательно, идет обмен аллелями генов.
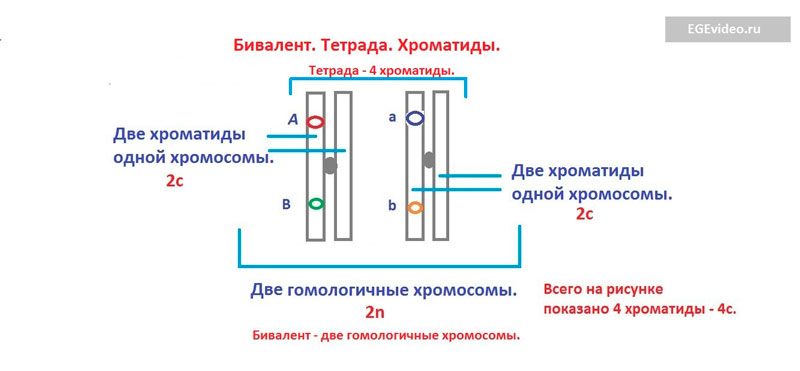
Что такое бивалент, тетрада?
Как вы знаете, семья состоит минимум из двух человек. Представьте, что пара рук мужчины - это одна гомологичная хромосома, пара рук женщины - вторая. Если мужчина и женщина соединят свои руки, получится метафора двух хромосом в профазе 1. Аналогично образуется бивалент. Две гомологичные хромосомы сближаются в профазе 1 для кроссинговера. Бивалент - это две объединенные в профазе 1 мейоза 1 гомологичные хромосомы. Так как в двух гомологичных хромосомах всего имеются 4 хроматиды, бивалент еще называют тетрадой.
Рисунок 2
Какова метафора кроссинговера?
Представим себе, что встретились два человека, словно две хромосомы. Допустим, данных людей сближает то, что они художники, профессионалы в одной области. Так и две хромосомы одинаковы в том, что они гомологичные - достались нам одна от отца, другая от матери, имеют взаимно параллельные участки и аллельные гены. У гипотетических художников цель общения - обмен опытом, идеями в изобразительном искусстве. Цель «общения» хромосом - обмен аллелями одного гена. Данные аллели (аллельные гены) близки тем, что они представляют один ген и отвечают за его альтернативные проявления. Например, рассмотрим ген цвета глаз. В каждой из гомологичных хромосом могут находиться по одному аллелю данного гена. Один аллель отвечает за карий цвет глаз, другой - за голубой.
После обмена идеями приобретут ли два художника новую профессию, например, инженера? Почему кроссинговер не образует новые аллели генов?
Вряд ли два наших художника изменят своему призванию. Так и гомологичные хромосомы после обмена не получат друг от друга абсолютно новых аллелей, например, аллель фиолетовых глаз. Они просто обменяются тем, что имеют. Если в одной хромосоме был аллель голубых глаз, она его передаст другой при кроссинговере. Гомологичная ей хромосома передаст свой ген карих глаз. В этом суть обмена. Сразу скажу, что совершенно новые аллели генов образуются в результате генных мутаций.
Рисунок 3. Отличия кроссинговера "до" и "после"






